Moss
20.34

There are approximately 12,000 species of moss classified in the Bryophyta.[2] The division Bryophyta formerly included not only mosses, but also liverworts and hornworts. These other two groups of bryophytes now are often placed in their own divisions.
Description
Botanically, mosses are bryophytes, or non-vascular plants. They can be distinguished from the apparently similar liverworts (Marchantiophyta or Hepaticae) by their multi-cellular rhizoids. Other differences are not universal for all mosses and all liverworts, but the presence of clearly differentiated "stem" and "leaves", the lack of deeply lobed or segmented leaves, and the absence of leaves arranged in three ranks, all point to the plant being a moss.
In addition to lacking a vascular system, mosses have a gametophyte-dominant life cycle, i.e. the plant's cells are haploid for most of its life cycle. Sporophytes (i.e. the diploid body) are short-lived and dependent on the gametophyte. This is in contrast to the pattern exhibited by most "higher" plants and by most animals. In seed plants, for example, the haploid generation is represented by the pollen and the ovule, whilst the diploid generation is the familiar flowering plant.
Life cycle
Most kinds of plants have two sets of chromosomes in their vegetative cells and are said to be diploid, i.e. each chromosome has a partner that contains the same, or similar, genetic information. By contrast, mosses and other bryophytes have only a single set of chromosomes and so are haploid (i.e. each chromosome exists in a unique copy within the cell). There are periods in the moss life cycle when they do have a double set of paired chromosomes, but this happens only during the sporophyte stage.
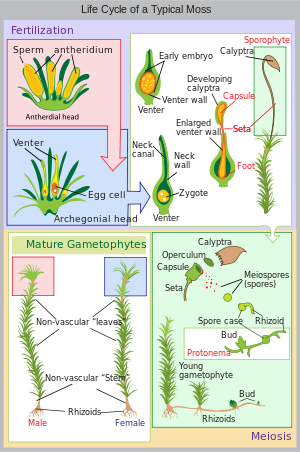
The life of a moss starts from a haploid spore. The spore germinates to produce a protonema (pl. protonemata), which is either a mass of thread-like filaments or thalloid (flat and thallus-like). Moss protonemata typically look like a thin green felt, and may grow on damp soil, tree bark, rocks, concrete, or almost any other reasonably stable surface. This is a transitory stage in the life of a moss, but from the protonema grows the gametophore ("gamete-bearer") that is structurally differentiated into stems and leaves. A single mat of protonemata may develop several gametophore shoots, resulting in a clump of moss.
From the tips of the gametophore stems or branches develop the sex organs of the mosses. The female organs are known as archegonia (sing. archegonium) and are protected by a group of modified leaves known as the perichaetum (plural, perichaeta). The archegonia are small flask-shaped clumps of cells with an open neck (venter) down which the male sperm swim. The male organs are known as antheridia (sing. antheridium) and are enclosed by modified leaves called the perigonium (pl. perigonia). The surrounding leaves in some mosses form a splash cup, allowing the sperm contained in the cup to be splashed to neighboring stalks by falling water droplets.
Mosses can be either dioicous (compare dioecious in seed plants) or monoicous (compare monoecious). In dioicous mosses, male and female sex organs are borne on different gametophyte plants. In monoicous (also called autoicous) mosses, both are borne on the same plant. In the presence of water, sperm from the antheridia swim to the archegonia and fertilisation occurs, leading to the production of a diploid sporophyte. The sperm of mosses is biflagellate, i.e. they have two flagellae that aid in propulsion. Since the sperm must swim to the archegonium, fertilisation cannot occur without water. After fertilisation, the immature sporophyte pushes its way out of the archegonial venter. It takes about a quarter to half a year for the sporophyte to mature. The sporophyte body comprises a long stalk, called a seta, and a capsule capped by a cap called the operculum. The capsule and operculum are in turn sheathed by a haploid calyptra which is the remains of the archegonial venter. The calyptra usually falls off when the capsule is mature. Within the capsule, spore-producing cells undergo meiosis to form haploid spores, upon which the cycle can start again. The mouth of the capsule is usually ringed by a set of teeth called peristome. This may be absent in some mosses.
In some mosses, e.g. Ulota phyllantha, green vegetative structures called gemmae are produced on leaves or branches, which can break off and form new plants without the need to go through the cycle of fertilization. This is a means of asexual reproduction, and the genetically identical units can lead to the formation of clonal populations.
Classification
Traditionally, mosses were grouped with the liverworts and hornworts in the Division Bryophyta (bryophytes), within which the mosses made up the class Musci. This definition of Bryophyta, however, is paraphyletic and now tends to be split up into three divisions. In such a system, the Division Bryophyta contains exclusively mosses.
The mosses are grouped as a single division, now named Bryophyta, and divided into eight classes:
division Bryophyta
|
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| The current phylogeny and composition of the Bryophyta.[2][3] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

Six of the eight classes contain only one or two genera each. Polytrichopsida includes 23 genera, and Bryopsida includes the majority of moss diversity with over 95% of moss species belonging to this class.
The Sphagnopsida, the peat-mosses, comprise the two living genera Ambuchanania and Sphagnum, as well as fossil taxa. However, the genus Sphagnum is a diverse, widespread, and economically important one. These large mosses form extensive acidic bogs in peat swamps. The leaves of Sphagnum have large dead cells alternating with living photosynthetic cells. The dead cells help to store water. Aside from this character, the unique branching, thallose (flat and expanded) protonema, and explosively rupturing sporangium place it apart from other mosses.
Andreaeopsida and Andreaeobryopsida are distinguished by the biseriate (two rows of cells) rhizoids, multiseriate (many rows of cells) protonema, and sporangium that splits along longitudinal lines. Most mosses have capsules that open at the top.
Polytrichopsida have leaves with sets of parallel lamellae, flaps of chloroplast-containing cells that look like the fins on a heat sink. These carry out photosynthesis and may help to conserve moisture by partially enclosing the gas exchange surfaces. The Polytrichopsida differ from other mosses in other details of their development and anatomy too, and can also become larger than most other mosses, with e.g. Polytrichum commune forming cushions up to 40 cm (16 in) high. The tallest land moss, a member of the Polytrichidae is probably Dawsonia superba, a native to New Zealand and other parts of Australasia.
They appear to be the closest living relatives of the vascular plants.

Geological history
The fossil record of moss is sparse, due to their soft-walled and fragile nature. Unambiguous moss fossils have been recovered from as early as the Permian of Antarctica and Russia, and a case is put forwards for Carboniferous mosses.[4] It has further been claimed that tube-like fossils from the Silurian are the macerated remains of moss calyptræ.[5]
Habitat





Mosses are found chiefly in areas of dampness and low light. Mosses are common in wooded areas and at the edges of streams. Mosses are also found in cracks between paving stones in damp city streets. Some types have adapted to urban conditions and are found only in cities. A few species are wholly aquatic, such as Fontinalis antipyretica, and others such as Sphagnum inhabit bogs, marshes and very slow-moving waterways. Such aquatic or semi-aquatic mosses can greatly exceed the normal range of lengths seen in terrestrial mosses. Individual plants 20–30 cm (8-12 in) or more long are common in Sphagnum species for example.
Wherever they occur, mosses require moisture to survive because of the small size and thinness of tissues, lack of cuticle (waxy covering to prevent water loss), and the need for liquid water to complete fertilisation. Some mosses can survive desiccation, returning to life within a few hours of rehydration.
In northern latitudes, the north side of trees and rocks will generally have more moss on average than other sides (though south-side outcroppings are not unknown). This is assumed to be because of the lack of sufficient water for reproduction on the sun-facing side of trees. South of the equator the reverse is true. In deep forests where sunlight does not penetrate, mosses grow equally well on all sides of the tree trunk.[citation needed]
Cultivation
Moss is considered a weed in grass lawns, but is deliberately encouraged to grow under aesthetic principles exemplified by Japanese gardening. In old temple gardens, moss can carpet a forest scene. Moss is thought to add a sense of calm, age, and stillness to a garden scene. Rules of cultivation are not widely established. Moss collections are quite often begun using samples transplanted from the wild in a water-retaining bag. However, specific species of moss can be extremely difficult to maintain away from their natural sites with their unique combinations of light, humidity, shelter from wind, etc.
Growing moss from spores is even less controlled. Moss spores fall in a constant rain on exposed surfaces; those surfaces which are hospitable to a certain species of moss will typically be colonised by that moss within a few years of exposure to wind and rain. Materials which are porous and moisture retentive, such as brick, wood, and certain coarse concrete mixtures are hospitable to moss. Surfaces can also be prepared with acidic substances, including buttermilk, yogurt, urine, and gently puréed mixtures of moss samples, water and ericaceous compost.
Inhibiting moss growth
Moss growth can be inhibited by a number of methods:
- Decreasing availability of water through drainage or direct application changes.
- Increasing direct sunlight.
- Increasing number and resources available for competitive plants like grasses.
- Increasing the soil pH with the application of lime.
Heavy traffic or manually disturbing the moss bed with a rake will also inhibit moss growth.
The application of products containing ferrous sulfate or ferrous ammonium sulfate will kill moss, these ingredients are typically in commercial moss control products and fertilizers. Sulfur and Iron are essential nutrients for some competing plants like grasses. Killing moss will not prevent regrowth unless conditions favorable to their growth are changed.[6]
Mossery
A passing fad for moss-collecting in the late 19th century led to the establishment of mosseries in many British and American gardens. The mossery is typically constructed out of slatted wood, with a flat roof, open to the north side (maintaining shade). Samples of moss were installed in the cracks between wood slats. The whole mossery would then be regularly moistened to maintain growth.
Commercial use
There is a substantial market in mosses gathered from the wild. The uses for intact moss are principally in the florist trade and for home decoration. Decaying moss in the genus Sphagnum is also the major component of peat, which is "mined" for use as a fuel, as a horticultural soil additive, and in smoking malt in the production of Scotch whisky.
Sphagnum moss, generally the species cristatum and subnitens, is harvested while still growing and is dried out to be used in nurseries and horticulture as a plant growing medium. The practice of harvesting peat moss should not be confused with the harvesting of moss peat.
Peat moss can be harvested on a sustainable basis and managed so that regrowth is allowed, whereas the harvesting of moss peat is generally considered to cause significant environmental damage as the peat is stripped with little or no chance of recovery.
In World War II, Sphagnum mosses were used as first-aid dressings on soldiers' wounds, as these mosses are highly absorbent and have mild antibacterial properties. Some early people used it as a diaper due to its high absorbency.[citation needed]
In rural UK, Fontinalis antipyretica was traditionally used to extinguish fires as it could be found in substantial quantities in slow-moving rivers and the moss retained large volumes of water which helped extinguish the flames. This historical use is reflected in its specific Latin/Greek name, the approximate meaning of which is "against fire".
In Finland, peat mosses have been used to make bread during famines.[citation needed]
In Mexico, Moss is used as a Christmas decoration.

Physcomitrella patens is increasingly used in biotechnology. Prominent examples are the identification of moss genes with implications for crop improvement or human health [7] and the safe production of complex biopharmaceuticals in the moss bioreactor, developed by Ralf Reski and his co-workers [8].


Posting Komentar