Atomic Spectra - 'fingerprints' for elements
21.04
Electrons exist in energy states within the atom (called orbitals by chemists).
Generally, the further away from the nucleus these states are, the higher the potential energy of the electron in that state. When an electron is 'free' of the nucleus' influence it is said to have zero potential energy - a zero energy state. When it is closer to the nucleus it has less potential energy (needs energy input to escape the influence). It is therefore at a negative energy level
We can put information about these energy states into a diagram

When an electron jumps between energy levels of an atom it either absorbs or emits a photon. The size of that photon's energy depends on the energy gap between the levels. This is linked to its frequency by the equation
E = hf
Where
h = Planck constant
f = frequency of the photon of electromagnetic radiation
(linked to its wavelength by c = f![]() )
)
Exciting the electrons
This can be done by bathing the gas sample in electromagnetic energy of a wide continuous spectrum (usually of visible light - but it is the same principle for other sections of the electromagnetic spectrum - only then you wouldn't 'see' the result - you would need to use a detector). The electrons absorb the photons they need to make transitions to higher energy levels and then give them back out again when they return to the ground state. They only remain in an excited state for less than a microsecond and are therefore constantly absorbing/emitting photons from the electromagnetic source.
There are two type of atomic spectra. The graphic below shows the visible pectra for hydrogen. You can see that the lines are in the same place on both spectra - because they correspond to the same energy jumps.


Photons that do not have energies that correspond to the 'gaps' between energy levels for the electrons are not absorbed. They go through the gas onto the photographic plate (or other form of detector).
When they give out the photons they have absorbed they do so in random directions therefore only a tiny fraction of them are emitted in the direction they would have gone had they not been absorbed in the frst place. This results in a lower intensity of electromagnetic radiation of the wavelength that has been absorbed by the sample shining onto a photographic plate. We therefore get an absorption spectrum
Click here for an interactive activity page on absoption spectra.

Another way of exciting the electrons is electrically. By putting the gas in a discharge tube we can 'zap' the atoms and therefore their electrons with electrical energy. This promotes electrons from the ground state to excited states and then as they 'jump down' they emit the characteristic photons of the jumps within the electron orbital of the atom. These can be collected by a dispersion system such as a spectrometer (or prism) which will disperse them into wavelengths.... missing wavelengths will not appear on the film, gaps will be left. The only coloured lines will be those that correspond to the wavelengths of the photons that 'fit' the energy jumps between orbitals. The spectrum we see is called an emission spectrum.
Click here for an interactive activity page on emission spectra.
We can look at these spectra using a spectrometer. This is an instrument that used either prisms to disperse the light by refraction or diffraction gratings to disperse the light by diffraction
Check out this link and look at the spectra of various gases in dischage tubes
The Atom
20.57
An atom is the smallest chemically indivisible part of an element - that means you can't split it by chemical (or physical) means into anything smaller - it takes a nuclear reaction to do it!
All of the different types of atom known to exist in the universe are listed in the periodic table.
Here is a simple diagram of the atom - you should KNOW this... including the sizes! Click here to find out how we know that the atom is like this.
At KS4 you do not need to know the mass of the particles in the atom in kiliogrammes. You just need to be able to compare the masses - to know that the mass of the proton is roughly the same as that of the neutron but that you would need almost 2000 electrons to have the same mass of electrons as that of a proton or neutron!

The above diagram is not to scale. If it was drawn to scale and you had a nucleus of 1 cm diameter in the centre of your screen you would need a screen of 1km diameter to show the full graphic! Most of the atom is empty space!.
For Chemists among you here is an interactive illustration of the orbital shapes of electrons.
Click here to find out more about the nucleus and click here for an overview of the atom.
Particle Accelerators
20.53
By accelerating particles to very high energies and smashing them into targets, or into each other physicists can unravel the forces acting between them.
Accelerators come in two types, linear (see the LINAC) and circular. Accelerators use powerful electric fields to push energy into a beam of particles - causing an increase in velocity and kinetic energy. Magnetic fields are used to keep the beam tightly focused, and in circular machines to steer the particles around the ring. See the cyclotron and the synchrotron.
Linear machines push energy into the beam all along the accelerator's length. The longer the machine, the higher the final energy, but this has practical implications!
 In circular machines the particles go round and round again, collecting energy with each lap. But the faster the particles are going, the more they try to 'skid' off the ring, just like cars going round a tight curve in the road. CERN's biggest accelerator, the Large Electron Positron collider LEP, is 27 kilometres round, keeping the curves as gentle as possible.
In circular machines the particles go round and round again, collecting energy with each lap. But the faster the particles are going, the more they try to 'skid' off the ring, just like cars going round a tight curve in the road. CERN's biggest accelerator, the Large Electron Positron collider LEP, is 27 kilometres round, keeping the curves as gentle as possible.
Atmospheric Pressure
20.39
|
At sea level the Earth's atmosphere is pushing on you from all sides with a pressure of 101,000 N/m2 (101 kPa). As you climb higher - away from sea level - that pressure decreases. This is because there is less air above you, weighing you down. We are not aware of the air pressure - we are designed to live with it. If we go to high altitudes we have difficulty breathing. This is because our lungs are used to taking in enough air to obtain the amount of oxygen we need. When we are at a high altitude a lung full of air contains less oxygen than it would on the ground because the density of the air is lower.We therefore breathe more rapidly at high altitudes until our bodies accustom themselves to the lower oxygen concentration. Weather and air pressure Atmospheric presure varies widely over the Earth's surface. It varies according to the amount of water vapour in the atmosphere in a locality and the temperature of the air molecules. This variation iscaused by uneven heating of the atmosphere by the Sun's rays. The pressure differences cause the winds to blow. Air moves from high pressure areas to low pressure areas. It does this most efficiently in a circular manner (like emptying a filled milk bottle is fastest if you swirl the fluid!). The weather stations map out the pressure and lines of equal pressure (isobars) on weather charts. They then use these to forcast the weather.
High pressure systems (anticyclones) are associated with settled, clear, cool, fresh weather. Around high-pressure systems, winds flow clockwise in the northern hemisphere, counterclockwise in the southern hemisphere.High pressure usually means good weather. Low pressure sytems (a depression) are generally unsettled and stormy due to the fact the warm air rises making the system it is on having low pressure. Which is generally the reason for storms and tropical cyclones.Tornadoes invariantly have very strong local low-pressure systems at their centres. Winds circulate around low pressure in an anti clockwise direction in the Northern Hemisphere.
|


Oceans
20.39
As water is much denser than air, this pressure increases as we venture into the water. For every 10m we decend into the depths of the ocean one more atmosphere of pressure pushes down on us. For example, at 20m deep we have three atmospheres of pressure pushing on us - two from the water and one from the air pushing on the sea's surface.

To travel into this high-pressure environment we have to make some adjustments. Humans can stand three or four atmospheres and still be okay. To go farther down we need to decend in a submarine.
Animals that live in the watery environment of the oceans undergo large pressure changes in short amounts of time - something that would give us 'the bends'. Sperm whales make hour-long dives 2,250 meters down. This is a pressure change of more than 223 atmospheres! By studying and understanding how these animals are able to withstand great pressure changes, scientists will be able to build better tools for humans to make such journeys.
Tides
Moss
20.34

There are approximately 12,000 species of moss classified in the Bryophyta.[2] The division Bryophyta formerly included not only mosses, but also liverworts and hornworts. These other two groups of bryophytes now are often placed in their own divisions.
Description
Botanically, mosses are bryophytes, or non-vascular plants. They can be distinguished from the apparently similar liverworts (Marchantiophyta or Hepaticae) by their multi-cellular rhizoids. Other differences are not universal for all mosses and all liverworts, but the presence of clearly differentiated "stem" and "leaves", the lack of deeply lobed or segmented leaves, and the absence of leaves arranged in three ranks, all point to the plant being a moss.
In addition to lacking a vascular system, mosses have a gametophyte-dominant life cycle, i.e. the plant's cells are haploid for most of its life cycle. Sporophytes (i.e. the diploid body) are short-lived and dependent on the gametophyte. This is in contrast to the pattern exhibited by most "higher" plants and by most animals. In seed plants, for example, the haploid generation is represented by the pollen and the ovule, whilst the diploid generation is the familiar flowering plant.
Life cycle
Most kinds of plants have two sets of chromosomes in their vegetative cells and are said to be diploid, i.e. each chromosome has a partner that contains the same, or similar, genetic information. By contrast, mosses and other bryophytes have only a single set of chromosomes and so are haploid (i.e. each chromosome exists in a unique copy within the cell). There are periods in the moss life cycle when they do have a double set of paired chromosomes, but this happens only during the sporophyte stage.
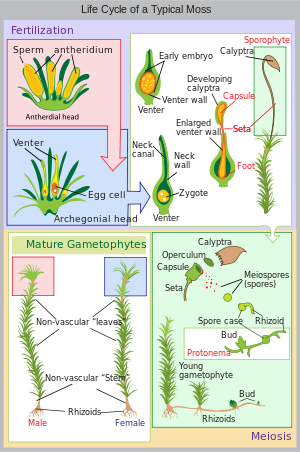
The life of a moss starts from a haploid spore. The spore germinates to produce a protonema (pl. protonemata), which is either a mass of thread-like filaments or thalloid (flat and thallus-like). Moss protonemata typically look like a thin green felt, and may grow on damp soil, tree bark, rocks, concrete, or almost any other reasonably stable surface. This is a transitory stage in the life of a moss, but from the protonema grows the gametophore ("gamete-bearer") that is structurally differentiated into stems and leaves. A single mat of protonemata may develop several gametophore shoots, resulting in a clump of moss.
From the tips of the gametophore stems or branches develop the sex organs of the mosses. The female organs are known as archegonia (sing. archegonium) and are protected by a group of modified leaves known as the perichaetum (plural, perichaeta). The archegonia are small flask-shaped clumps of cells with an open neck (venter) down which the male sperm swim. The male organs are known as antheridia (sing. antheridium) and are enclosed by modified leaves called the perigonium (pl. perigonia). The surrounding leaves in some mosses form a splash cup, allowing the sperm contained in the cup to be splashed to neighboring stalks by falling water droplets.
Mosses can be either dioicous (compare dioecious in seed plants) or monoicous (compare monoecious). In dioicous mosses, male and female sex organs are borne on different gametophyte plants. In monoicous (also called autoicous) mosses, both are borne on the same plant. In the presence of water, sperm from the antheridia swim to the archegonia and fertilisation occurs, leading to the production of a diploid sporophyte. The sperm of mosses is biflagellate, i.e. they have two flagellae that aid in propulsion. Since the sperm must swim to the archegonium, fertilisation cannot occur without water. After fertilisation, the immature sporophyte pushes its way out of the archegonial venter. It takes about a quarter to half a year for the sporophyte to mature. The sporophyte body comprises a long stalk, called a seta, and a capsule capped by a cap called the operculum. The capsule and operculum are in turn sheathed by a haploid calyptra which is the remains of the archegonial venter. The calyptra usually falls off when the capsule is mature. Within the capsule, spore-producing cells undergo meiosis to form haploid spores, upon which the cycle can start again. The mouth of the capsule is usually ringed by a set of teeth called peristome. This may be absent in some mosses.
In some mosses, e.g. Ulota phyllantha, green vegetative structures called gemmae are produced on leaves or branches, which can break off and form new plants without the need to go through the cycle of fertilization. This is a means of asexual reproduction, and the genetically identical units can lead to the formation of clonal populations.
Classification
Traditionally, mosses were grouped with the liverworts and hornworts in the Division Bryophyta (bryophytes), within which the mosses made up the class Musci. This definition of Bryophyta, however, is paraphyletic and now tends to be split up into three divisions. In such a system, the Division Bryophyta contains exclusively mosses.
The mosses are grouped as a single division, now named Bryophyta, and divided into eight classes:
division Bryophyta
|
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| The current phylogeny and composition of the Bryophyta.[2][3] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

Six of the eight classes contain only one or two genera each. Polytrichopsida includes 23 genera, and Bryopsida includes the majority of moss diversity with over 95% of moss species belonging to this class.
The Sphagnopsida, the peat-mosses, comprise the two living genera Ambuchanania and Sphagnum, as well as fossil taxa. However, the genus Sphagnum is a diverse, widespread, and economically important one. These large mosses form extensive acidic bogs in peat swamps. The leaves of Sphagnum have large dead cells alternating with living photosynthetic cells. The dead cells help to store water. Aside from this character, the unique branching, thallose (flat and expanded) protonema, and explosively rupturing sporangium place it apart from other mosses.
Andreaeopsida and Andreaeobryopsida are distinguished by the biseriate (two rows of cells) rhizoids, multiseriate (many rows of cells) protonema, and sporangium that splits along longitudinal lines. Most mosses have capsules that open at the top.
Polytrichopsida have leaves with sets of parallel lamellae, flaps of chloroplast-containing cells that look like the fins on a heat sink. These carry out photosynthesis and may help to conserve moisture by partially enclosing the gas exchange surfaces. The Polytrichopsida differ from other mosses in other details of their development and anatomy too, and can also become larger than most other mosses, with e.g. Polytrichum commune forming cushions up to 40 cm (16 in) high. The tallest land moss, a member of the Polytrichidae is probably Dawsonia superba, a native to New Zealand and other parts of Australasia.
They appear to be the closest living relatives of the vascular plants.

Geological history
The fossil record of moss is sparse, due to their soft-walled and fragile nature. Unambiguous moss fossils have been recovered from as early as the Permian of Antarctica and Russia, and a case is put forwards for Carboniferous mosses.[4] It has further been claimed that tube-like fossils from the Silurian are the macerated remains of moss calyptræ.[5]
Habitat





Mosses are found chiefly in areas of dampness and low light. Mosses are common in wooded areas and at the edges of streams. Mosses are also found in cracks between paving stones in damp city streets. Some types have adapted to urban conditions and are found only in cities. A few species are wholly aquatic, such as Fontinalis antipyretica, and others such as Sphagnum inhabit bogs, marshes and very slow-moving waterways. Such aquatic or semi-aquatic mosses can greatly exceed the normal range of lengths seen in terrestrial mosses. Individual plants 20–30 cm (8-12 in) or more long are common in Sphagnum species for example.
Wherever they occur, mosses require moisture to survive because of the small size and thinness of tissues, lack of cuticle (waxy covering to prevent water loss), and the need for liquid water to complete fertilisation. Some mosses can survive desiccation, returning to life within a few hours of rehydration.
In northern latitudes, the north side of trees and rocks will generally have more moss on average than other sides (though south-side outcroppings are not unknown). This is assumed to be because of the lack of sufficient water for reproduction on the sun-facing side of trees. South of the equator the reverse is true. In deep forests where sunlight does not penetrate, mosses grow equally well on all sides of the tree trunk.[citation needed]
Cultivation
Moss is considered a weed in grass lawns, but is deliberately encouraged to grow under aesthetic principles exemplified by Japanese gardening. In old temple gardens, moss can carpet a forest scene. Moss is thought to add a sense of calm, age, and stillness to a garden scene. Rules of cultivation are not widely established. Moss collections are quite often begun using samples transplanted from the wild in a water-retaining bag. However, specific species of moss can be extremely difficult to maintain away from their natural sites with their unique combinations of light, humidity, shelter from wind, etc.
Growing moss from spores is even less controlled. Moss spores fall in a constant rain on exposed surfaces; those surfaces which are hospitable to a certain species of moss will typically be colonised by that moss within a few years of exposure to wind and rain. Materials which are porous and moisture retentive, such as brick, wood, and certain coarse concrete mixtures are hospitable to moss. Surfaces can also be prepared with acidic substances, including buttermilk, yogurt, urine, and gently puréed mixtures of moss samples, water and ericaceous compost.
Inhibiting moss growth
Moss growth can be inhibited by a number of methods:
- Decreasing availability of water through drainage or direct application changes.
- Increasing direct sunlight.
- Increasing number and resources available for competitive plants like grasses.
- Increasing the soil pH with the application of lime.
Heavy traffic or manually disturbing the moss bed with a rake will also inhibit moss growth.
The application of products containing ferrous sulfate or ferrous ammonium sulfate will kill moss, these ingredients are typically in commercial moss control products and fertilizers. Sulfur and Iron are essential nutrients for some competing plants like grasses. Killing moss will not prevent regrowth unless conditions favorable to their growth are changed.[6]
Mossery
A passing fad for moss-collecting in the late 19th century led to the establishment of mosseries in many British and American gardens. The mossery is typically constructed out of slatted wood, with a flat roof, open to the north side (maintaining shade). Samples of moss were installed in the cracks between wood slats. The whole mossery would then be regularly moistened to maintain growth.
Commercial use
There is a substantial market in mosses gathered from the wild. The uses for intact moss are principally in the florist trade and for home decoration. Decaying moss in the genus Sphagnum is also the major component of peat, which is "mined" for use as a fuel, as a horticultural soil additive, and in smoking malt in the production of Scotch whisky.
Sphagnum moss, generally the species cristatum and subnitens, is harvested while still growing and is dried out to be used in nurseries and horticulture as a plant growing medium. The practice of harvesting peat moss should not be confused with the harvesting of moss peat.
Peat moss can be harvested on a sustainable basis and managed so that regrowth is allowed, whereas the harvesting of moss peat is generally considered to cause significant environmental damage as the peat is stripped with little or no chance of recovery.
In World War II, Sphagnum mosses were used as first-aid dressings on soldiers' wounds, as these mosses are highly absorbent and have mild antibacterial properties. Some early people used it as a diaper due to its high absorbency.[citation needed]
In rural UK, Fontinalis antipyretica was traditionally used to extinguish fires as it could be found in substantial quantities in slow-moving rivers and the moss retained large volumes of water which helped extinguish the flames. This historical use is reflected in its specific Latin/Greek name, the approximate meaning of which is "against fire".
In Finland, peat mosses have been used to make bread during famines.[citation needed]
In Mexico, Moss is used as a Christmas decoration.

Physcomitrella patens is increasingly used in biotechnology. Prominent examples are the identification of moss genes with implications for crop improvement or human health [7] and the safe production of complex biopharmaceuticals in the moss bioreactor, developed by Ralf Reski and his co-workers [8].
Hornwort
20.30

Hornworts are a group of bryophytes, or non-vascular plants, comprising the division Anthocerotophyta. The common name refers to the elongated horn-like structure, which is the sporophyte. The flattened, green plant body of a hornwort is the gametophyte plant.
Hornworts may be found world-wide, though they tend to grow only in places that are damp or humid. Some species grow in large numbers as tiny weeds in the soil of gardens and cultivated fields. Large tropical and sub-tropical species of Dendroceros may be found growing on the bark of trees.
Description
The plant body of a hornwort is a haploid gametophyte stage. This stage usually grows as a thin rosette or ribbon-like thallus between one and five centimeters in diameter. Each cell of the thallus usually contains just one chloroplast per cell. In most species, this chloroplast is fused with other organelles to form a large pyrenoid that both manufactures and stores food. This particular feature is very unusual in land plants, but is common among algae.
Many hornworts develop internal mucilage-filled cavities when groups of cells break down. These cavities are invaded by photosynthetic cyanobacteria, especially species of Nostoc. Such colonies of bacteria growing inside the thallus give the hornwort a distinctive blue-green color. There may also be small slime pores on the underside of the thallus. These pores superficially resemble the stomata of other plants.
The horn-shaped sporophyte grows from an archegonium embedded deep in the gametophyte. Hornworts sporophytes are unusual in that the sporophyte grows from a meristem near its base, instead of from its tip the way other plants do. Unlike liverworts, most hornworts have true stomata on the sporophyte as mosses do. The exceptions are the genera Notothylas and Megaceros, which do not have stomata.
When the sporophyte is mature, it has a multicellular outer layer, a central rod-like columella running up the center, and a layer of tissue in between that produces spores and pseudo-elaters. The pseudo-elaters are multi-cellular, unlike the elaters of liverworts. They have helical thickenings that change shape in response to drying out; they twist and thereby help to disperse the spores. Hornwort spores are relatively large for bryophytes, measuring between 30 and 80 µm in diameter or more. The spores are polar, usually with a distinctive Y-shaped tri-radiate ridge on the proximal surface, and with a distal surface ornamented with bumps or spines.
Life cycle
The life of a hornwort starts from a haploid spore. In most species, there is a single cell inside the spore, and a slender extension of this cell called the germ tube germinates from the proximal side of the spore. The tip of the germ tube divides to form an octant of cells, and the first rhizoid grows as an extension of the original germ cell. The tip continues to divide new cells, which produces a thalloid protonema. By contrast, species of the family Dendrocerotaceae may begin dividing within the spore, becoming multicellular and even photosynthetic before the spore germinates. In either case, the protonema is a transitory stage in the life of a hornwort.

From the protonema grows the adult gametophyte, which is the persistent and independent stage in the life cycle. This stage usually grows as a thin rosette or ribbon-like thallus between one and five centimeters in diameter, and several layers of cells in thickness. It is green or yellow-green from the chlorophyll in its cells, or bluish-green when colonies of cyanobacteria grow inside the plant.
When the gametophyte has grown to its adult size, it produces the sex organs of the hornwort. Most plants are monoicous, with both sex organs on the same plant, but some plants (even within the same species) are dioicous, with separate male and female gametophytes. The female organs are known as archegonia (singular archegonium) and the male organs are known as antheridia (singular antheridium). Both kinds of organs develop just below the surface of the plant and are only later exposed by disintegration of the overlying cells.
The biflagellate sperm must swim from the antheridia, or else be splashed to the archegonia. When this happens, the sperm and egg cell fuse to form a zygote, the cell from which the sporophyte stage of the life cycle will develop. Unlike all other bryophytes, the first cell division of the zygote is longitudinal. Further divisions produce three basic regions of the sporophyte.
At the bottom of the sporophyte (closest to the interior of the gametophyte), is a foot. This is a globular group of cells that receives nutrients from the parent gametophyte, on which the sporophyte will spend its entire existence. In the middle of the sporophyte (just above the foot), is a meristem that will continue to divide and produce new cells for the third region. This third region is the capsule. Both the central and surface cells of the capsule are sterile, but between them is a layer of cells that will divide to produce pseudo-elaters and spores. These are released from the capsule when it splits lengthwise from the tip.
Evolutionary history
While the fossil record of crown group hornworts only begins in the upper Cretaceous, the lower Devonian Horneophyton may represent a stem group to the clade, as it possesses a sporangium with central columella not attached at the roof.[2] However, the same form of columella is also characteristic of basal moss groups, such as the Sphagnopsida and Andreaeopsida, and has been interpreted as a character common to all early land plants with stomata.[3]
Classification of Hornworts
Hornworts were traditionally considered a class within the Division Bryophyta (bryophytes). However, it now appears that this group is paraphyletic, so the hornworts tend to be given their own division, called Anthocerotophyta. The Bryophyta is now restricted to include only mosses.
There is a single class of hornworts, called Anthocerotopsida, or traditionally Anthocerotae. This class includes a single order of hornworts (Anthocerotales) in this classification scheme. In some other classification schemes, a second order Notothyladales (containing only the genus Notothylas) is recognized because of the unique and unusual features present in that group.
Among land plants, hornworts appear to be one of the oldest surviving lineages; cladistic analysis implies that the group originated prior to the Devonian, around the same time as the mosses and liverworts. There are only about 100 species known, but new species are still being discovered. The number and names of genera are a current matter of investigation, and several competing classification schemes have been published since 1988.
A more recent study of molecular, ultrastructural and morphological data has yielded a new classification of hornworts,[4] given below:

Families and genera
Leiosporocerotaceae
- Leiosporoceros
Anthocerotaceae
- Anthoceros
- Folioceros
- Sphaerosporoceros
Notothyladaceae
- Notothylas
- Phaeoceros
- Paraphymatoceros
- Hattorioceros
- Mesoceros
Phymatocerotaceae
- Phymatoceros
Dendrocerotaceae
- Dendroceros
- Megaceros
- Nothoceros
- Phaeomegaceros
Marchantiophyta
20.26

It is estimated that there are 6000 to 8000 species of liverworts, though when Neotropical regions are better studied this number may approach 10,000. Some of the more familiar species grow as a flattened leafless thallus, but most species are leafy with a form very much like a flattened moss. Leafy species can be distinguished from the apparently similar mosses on the basis of a number of features, including their single-celled rhizoids. Leafy liverworts also differ from most (but not all) mosses in that their leaves never have a costa (present in many mosses) and may bear marginal cilia (very rare in mosses). Other differences are not universal for all mosses and liverworts, but the occurrence of leaves arranged in three ranks, the presence of deep lobes or segmented leaves, or a lack of clearly differentiated stem and leaves all point to the plant being a liverwort.
Physical characteristics
Description
Most liverworts are small, usually from 2–20 millimetres (0.08–0.8 in) wide with individual plants less than 10 centimetres (4 in) long,[5] so they are often overlooked. The most familiar liverworts consist of a prostrate, flattened, ribbon-like or branching structure called a thallus (plant body); these liverworts are termed thallose liverworts. However, most liverworts produce flattened stems with overlapping scales or leaves in three or more ranks, the middle rank being conspicuously different from the outer ranks; these are called leafy liverworts or scale liverworts.[6][7] (See the gallery below for examples.)

Liverworts can most reliably be distinguished from the apparently similar mosses by their single-celled rhizoids.[8] Other differences are not universal for all mosses and all liverworts;[7] but the lack of clearly differentiated stem and leaves in thallose species, or in leafy species the presence of deeply lobed or segmented leaves and the presence of leaves arranged in three ranks, all point to the plant being a liverwort.[9][10] In addition, 90% of liverworts contain oil bodies in at least some of their cells, and these cellular structures are absent from most other bryophytes and from all vascular plants.[11] The overall physical similarity of some mosses and leafy liverworts means that confirmation of the identification of some groups can be performed with certainty only with the aid of microscopy or an experienced bryologist.
Liverworts have a gametophyte-dominant life cycle, with the sporophyte dependent on the gametophyte.[11] Cells in a typical liverwort plant each contain only a single set of genetic information, so the plant's cells are haploid for the majority of its life cycle. This contrasts sharply with the pattern exhibited by nearly all animals and by most other plants. In the more familiar seed plants, the haploid generation is represented only by the tiny pollen and the ovule, while the diploid generation is the familiar tree or other plant.[12] Another unusual feature of the liverwort life cycle is that sporophytes (i.e. the diploid body) are very short-lived, withering away not long after releasing spores.[13] Even in other bryophytes, the sporophyte is persistent and disperses spores over an extended period.
Life cycle

The life of a liverwort starts from the germination of a haploid spore to produce a protonema, which is either a mass of thread-like filaments or else a flattened thallus.[14][15] The protonema is a transitory stage in the life of a liverwort, from which will grow the mature gametophore ("gamete-bearer") plant that produces the sex organs. The male organs are known as antheridia (singular: antheridium) and produce the sperm cells. Clusters of antheridia are enclosed by a protective layer of cells called the perigonium (plural: perigonia). As in other land plants, the female organs are known as archegonia (singular: archegonium) and are protected by the thin surrounding perichaetum (plural: perichaeta).[7] Each archegonium has a slender hollow tube, the "neck", down which the sperm swim to reach the egg cell.
Liverwort species may be either dioicous or monoicous. In dioicious liverworts, female and male sex organs are borne on different and separate gametophyte plants. In monoicious liverworts, the two kinds of reproductive structures are borne on different branches of the same plant.[16] In either case, the sperm must swim from the antheridia where they are produced to the archegonium where the eggs are held. The sperm of liverworts is biflagellate, i.e. they have two tail-like flagellae that aid in propulsion.[17] Their journey is further assisted either by the splashing of raindrops or the presence of a thin layer of water covering the plants. Without water, the journey from antheridium to archegonium cannot occur.
In the presence of such water, sperm from the antheridia swim to the archegonia and fertilisation occurs, leading to the production of a diploid sporophyte. After fertilisation, the immature sporophyte within the archegonium develops three distinct regions: (1) a foot, which both anchors the sporophyte in place and receives nutrients from its "mother" plant, (2) a spherical or ellipsoidal capsule, inside which the spores will be produced for dispersing to new locations, and (3) a seta (stalk) which lies between the other two regions and connects them.[17] When the sporophyte has developed all three regions, the seta elongates, pushing its way out of the archegonium and rupturing it. While the foot remains anchored within the parent plant, the capsule is forced out by the seta and is extended away from the plant and into the air. Within the capsule, cells divide to produce both elater cells and spore-producing cells. The elaters are spring-like, and will push open the wall of the capsule to scatter themselves when the capsule bursts. The spore-producing cells will undergo meiosis to form haploid spores to disperse, upon which point the life cycle can start again.
Ecology
Today, liverworts can be found in many ecosystems across the planet except the sea and excessively dry environments, or those exposed to high levels of direct solar radiation.[18] As with most groups of living plants, they are most common (both in numbers and species) in moist tropical areas.[19] Liverworts are more commonly found in moderate to deep shade, though desert species may tolerate direct sunlight and periods of total desiccation.
Classification
Relationship to other plants
Traditionally, the liverworts were grouped together with other bryophytes (mosses and hornworts) in the Division Bryophyta, within which the liverworts made up the class Hepaticae (also called Marchantiopsida).[20][7] However, since this grouping makes the Bryophyta paraphyletic, the liverworts are now usually given their own division.[21] The use of the division name Bryophyta sensu latu is still found in the literature, but more frequently the Bryophyta now is used in a restricted sense to include only the mosses.
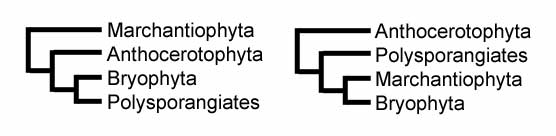
Another reason that liverworts are now classified separately is that they appear to have diverged from all other embryophyte plants near the beginning of their evolution. The strongest line of supporting evidence is that liverworts are the only living group of land plants that do not have stomata on the sporophyte generation.[22] Among the earliest fossils believed to be liverworts are compression fossils of Pallaviciniites from the Upper Devonian of New York.[23] These fossils resemble modern species in the Metzgeriales.[24] Another Devonian fossil called Protosalvinia also looks like a liverwort, but its relationship to other plants is still uncertain, so it may not belong to the Marchantiophyta. In 2007, the oldest fossils assignable to the liverworts were announced, Metzgeriothallus sharonae from the Givetian (Middle Devonian) of New York, USA.[1]
Internal classification
Bryologists classify liverworts in the division Marchantiophyta. This divisional name is based on the name of the most universally recognized liverwort genus Marchantia.[25] In addition to this taxon-based name, the liverworts are often called Hepaticophyta. This name is derived from their common Latin name as Latin was the language in which botanists published their descriptions of species. This name has led to some confusion, partly because it appears to be a taxon-based name derived from the genus Hepatica which is actually a flowering plant of the buttercup family Ranunculaceae. In addition, the name Hepaticophyta is frequently misspelled in textbooks as Hepatophyta, which only adds to the confusion.
Although there is no consensus among bryologists as to the classification of liverworts above family rank,[26] the Marchantiophyta may be subdivided into three classes:[27][28][29][30]
- The Jungermanniopsida includes the two orders Metzgeriales (simple thalloids) and Jungermanniales (leafy liverworts).
- The Marchantiopsida includes the three orders Marchantiales (complex-thallus liverworts), and Sphaerocarpales (bottle hepatics), as well as the Blasiales (previously placed among the Metzgeriales).[27][31] It also includes the problematic genus Monoclea, which is sometimes placed in its own order Monocleales.[32]
- A third class, the Haplomitriopsida is newly recognized as a basal sister group to the other liverworts;[30] it comprises the genera Haplomitrium, Treubia, and Apotreubia.
| The diagram at right summarizes a portion of a 2006 cladistic analysis of liverworts based upon three chloroplast genes, one nuclear gene, and one mitochondrial gene.[27] |
|
It is estimated that there are 6000 to 8000 species of liverworts, at least 85% of which belong to the leafy group.[3]
Economic importance
In ancient times, it was believed that liverworts cured diseases of the liver, hence the name.[33] In Old English, the word liverwort literally means liver plant.[34] This probably stemmed from the superficial appearance of some thalloid liverworts (which resemble a liver in outline), and led to the common name of the group as hepatics, from the Latin word hēpaticus for "belonging to the liver". An unrelated flowering plant, Hepatica, is sometimes also referred to as liverwort because it was once also used in treating diseases of the liver. This archaic relationship of plant form to function was based in the "Doctrine of Signatures".[35]
Liverworts have little direct economic importance today. Their greatest impact is indirect, through the reduction of erosion along streambanks, their collection and retention of water in tropical forests, and the formation of soil crusts in deserts and polar regions. However, a few species are used by humans directly. A few species, such as Riccia fluitans, are aquatic thallose liverworts sold for use in aquaria. Their thin, slender branches float on the water's surface and provide habitat for both small invertebrates and the fish that feed on them.

